
菌根について学ぼう Vol.2 “ラン型菌根ってなに?”
前回は菌根の概要について説明しました。今回からはラン型菌根の特徴について解説していきます(前回からかなり時間が空いてしまいました、すみません)。
この記事の目次
ラン科植物とは
まず、このページを見ている皆さんが大好きな、ラン科植物について軽く触れておきます。
ラン科は単子葉類のキジカクシ目に属する、約700属・28000種以上が知られているとても大きなグループです(Chase et al., 2015)。
これは被子植物全体の種数の約10%にも相当します。適当に植物の学名を挙げると、10種に1種はラン科植物ということですね。加えて、年平均500種の新種が記載されているそうで、まだまだ知られていない種が多く存在しています。
ラン科植物の多様性の高さは生息環境の多様さ、送粉様式の多様さや種特異的な送粉者との共生などに関係していると考えられます(Peakall, 2007)。
ラン科植物は熱帯から亜寒帯までの樹林下や草原、湿地や乾燥地帯にも生息し、地表のみではなく樹上にも進出しました。維管束着生植物の70%がラン科植物ということからも、その特殊さが分かります(Zotz et al., 2013)。
送粉様式に関しては、ラン科植物は送粉者を騙すことに長けた植物であると言えます。1/3ほどのランが送粉者に報酬(蜜や花粉など)を与えない、無報酬花をつけます(Peakall, 2007)。そこにエサがあると思い込ませ、送粉者をおびき寄せる作戦です。また、特定の昆虫の性フェロモン様物質を分泌し、虫を性的に騙すランもいます。虫を性的に騙すランはOphylis属やCaladenia属が有名です(Schiestl et al., 1999; Phillips et al., 2009)。
このように多様な生態を持ち、多様な花を作ることで、ランは人を虜にしたと言えるでしょう。
ラン科植物は以下の5亜科に分けられます(Chase et al., 2015)。
・ヤクシマラン亜科(ヤクシマラン属など)
・バニラ亜科(バニラ属やムヨウラン属、トキソウ属など)
・アツモリソウ亜科(アツモリソウ属やパフィオペディラム属など)
・ラン亜科(シュスラン属やネジバナ属など)
・セッコク亜科(カトレア属やデンドロビウム属、シュンラン属など)
ラン型菌根の特徴
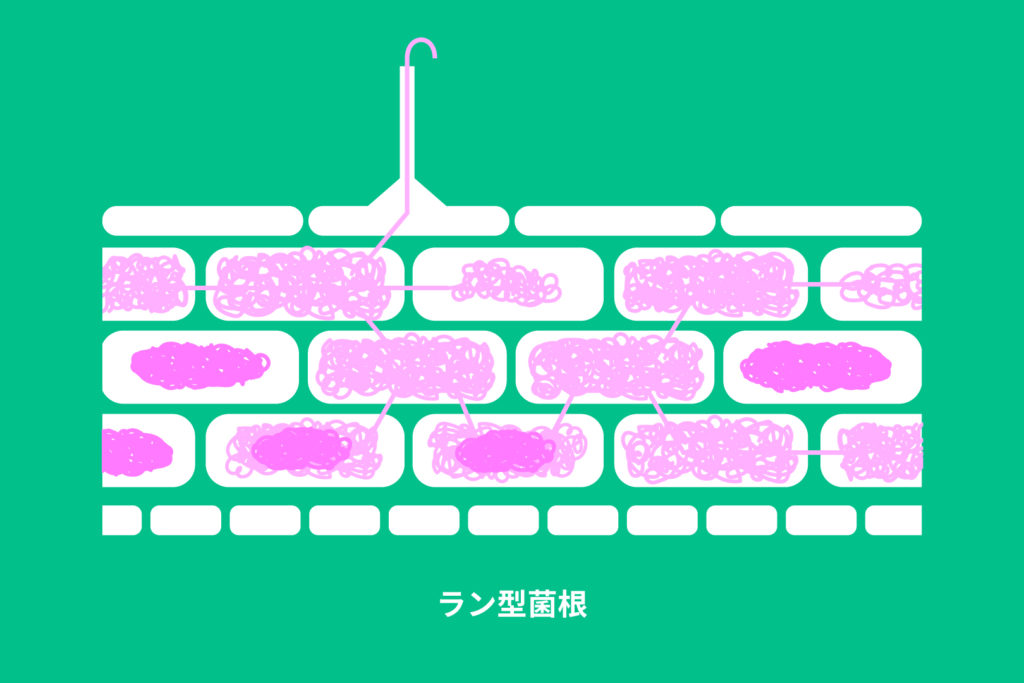
ラン型菌根とは、前回の記事でも説明した通り、ラン科植物が形成する菌根のことです。ラン科植物の中でも、最も原始的なヤクシマラン亜科を含む、全てのラン科植物がラン型菌根を形成します。
キジカクシ目の中では、ラン科植物以外はアーバスキュラー菌根を形成することから、ラン科植物の共通祖先において菌根の変化が起きたと考えられます(Smith and Read, 2008; Yukawa et al., 2009)。
ラン型菌根では、菌根菌は植物の皮層細胞内に、菌毬(Peloton)と呼ばれる菌糸コイルを形成します。菌毬は下の写真の様な形態をしています。菌糸は植物細胞膜内に侵入することはなく、植物細胞膜で覆われた状態になっており、この菌糸と植物細胞膜間の隙間を利用して、ランと菌は栄養の交換を行います。また、菌毬は形成されてから十数時間後から数日後には崩壊し始めます。この菌毬の崩壊のことを菌毬消化と呼びます。ランは菌毬が崩壊する際に多くの栄養を得ているようなので(Kuga et al., 2014)、菌を食べていると表現することがあります。
古くから、ラン型菌根では、菌根菌はランからほとんど利益を得ておらず、ランが菌に寄生している状態だと言われてきました。そのため、ラン型菌根は「ランと菌のバトルフィールド」と呼ばれることもあったようです(Smith and Read, 2008)。
現在では、菌根菌がランから炭素をもらうことがあると分かってきました(Cameron et al. 2008; Latalova and Balaz, 2010)。また、ランは菌の侵入・定着の際に防御反応を示していないことから、ランと菌の関係は友好的であると考えるのが定説となってきています(Dearnaley et al., 2016)。
しかし、独立栄養だと考えられてきたランも菌から炭素をもらっているという報告もあり(Girlanda et al., 2011; Gebauer et al., 2016)、ランと菌の間の炭素のやり取りに関してはまだ不透明なままと言えるでしょう。
一方で、確実にランが菌に依存していると考えられるような例も数多くあります。キンラン属やホテイランなどの部分的菌従属栄養植物や、マヤランやタシロラン等の完全菌従属栄養植物と菌根菌との共生がそれです。
前者の部分的菌従属栄養植物は、葉緑体を持ち光合成をしますが、自力で合成できる有機炭素のみでは生活が出来ず、菌類にも炭素を頼っている植物を指します。これらの菌従属栄養性のランは、主に樹木と共生する菌根菌である外生菌根菌(ベニタケ属やロウタケ属など)や、木を腐らせる菌である木材腐朽菌(ナヨタケ属やクヌギタケ属など)と共生しています(Dearnaley et al., 2012)。菌根菌は光合成産物の有機炭素をもらっていないか、もらっていてもランに与えている分の方が多いと考えられています。
ラン科には菌従属栄養植物が多く存在していますが、それは次に説明する種子発芽方式に関係していると考えられます。
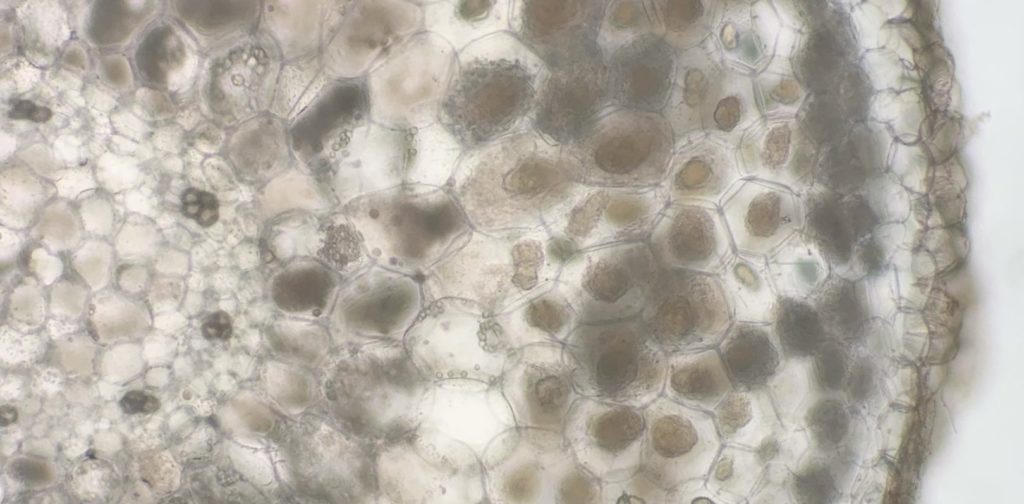
ナンゴクネジバナの根の横断切片。植物細胞内のモヤモヤしているものがアクティブな菌毬で、黄色っぽくなっているのが崩壊途中の菌毬。スケールを入れ忘れたが、ナンゴクネジバナの根の半径は1.5mmほど。
ランの種子発芽
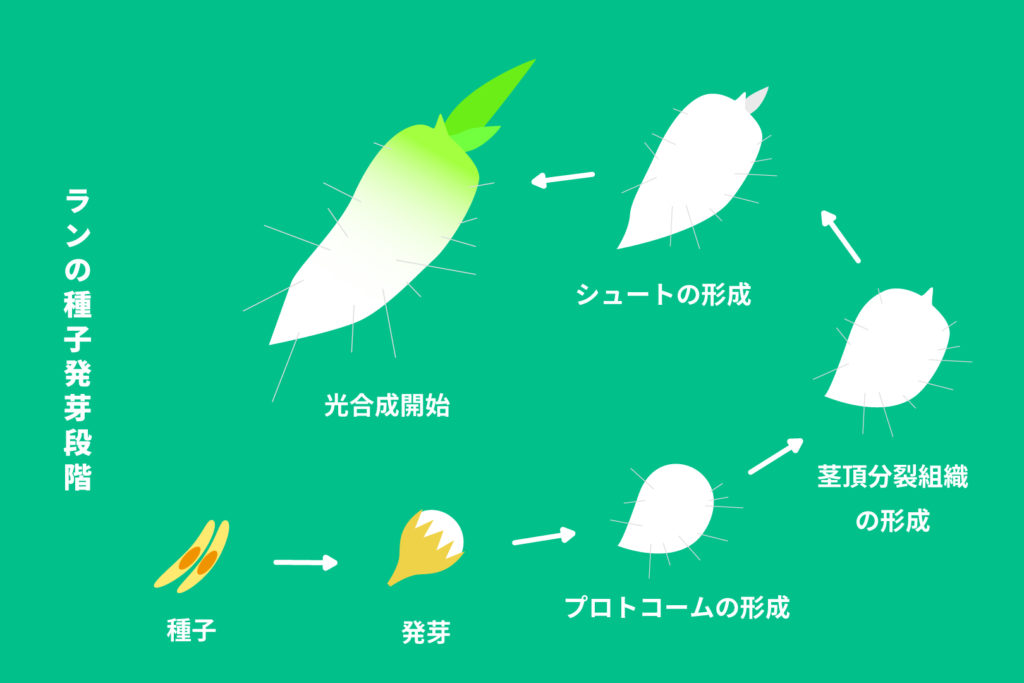
ラン科植物の種子は上図のような形態をしており、大きさは大きいものでも1㎜程度です。この種子は埃種子や微細種子と呼ばれ、非常に小さく、自力で発芽し成長するだけのエネルギーを持っていません。
ランの種子は発芽後、プロトコームを形成します。プロトコームはあまり組織の分化の無い細胞の塊で、ランはここでも菌根菌と共生します。菌と共生できたプロトコームはその後、茎頂分裂組織を分化させ、そこからシュート(芽)を出します。
ここまでの期間は光合成が出来ないため、ランは菌に完全に依存して成長することになります。このように、生活史の初期に菌に依存して生活することを生活史初期型菌従属栄養と呼び、菌と共生して発芽することを共生発芽と呼びます。
しかし、皆さんご存じのように、ランの種子は無菌でも発芽させることが出来ます。ランの種子は糖類やビタミン等を成長のために必要としており、それらを人工的に供給してあげることで、発芽させることが出来ます。自然界には、ランの発芽から自力で光合成できるようになるまでの成長を支えられるだけの糖類が存在することはほぼあり得ないため、自然界での種子発芽には菌根菌の存在が必要不可欠なのです(Smith and Read, 2008)。
プロトコームでも、菌根菌はランの細胞内に菌毬を形成します。菌根の構造や共生する菌のグループはプロトコームも根も違いはなく、プロトコームの共生菌も菌根菌と呼ばれることがあります。
種子のサイズと生産数はトレードオフの関係にあります。種子のサイズを大きくすると、外的ストレスに対する耐久性が高くなり、生産数を大きくすると種子が発芽に適したニッチに到達できる可能性が上がります。
ラン科植物は菌との共生を発達させたことで、種子のサイズを極限まで小さくすることに成功しました。そして種子生産数と散布効率を上げることが出来ました。ラン科植物が陸上植物の中で最も多くの種数を占めるということは、この戦略は一定の成功を修めたと言えるでしょう(Eriksson and Kainulainen, 2011)。
おわりに
今回はラン型菌根の構造やランと菌の関係性について解説しました。次回はランの大事なパートナーである菌根菌の種類と、1種のランが何種の菌根菌と共生できるのかを表す、菌根特異性について解説する予定です。
また時間がかかってしまうかもしれませんが、気長に待っていただけるとありがたいです。
引用文献
Smith, S. E. & Read, D. J. (2008). Mycorrhizal symbiosis. Academic Press, Cambridge, UK.


